
実験動物のより良い未来を模索する
実験動物のより良い未来を模索する



- 2026.07.23
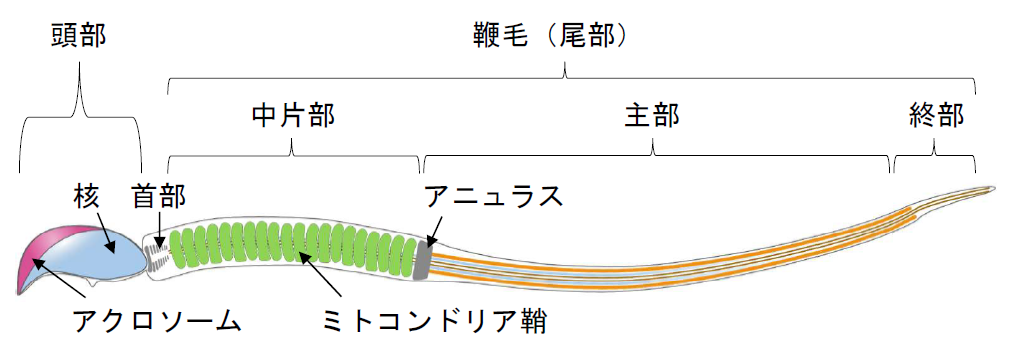
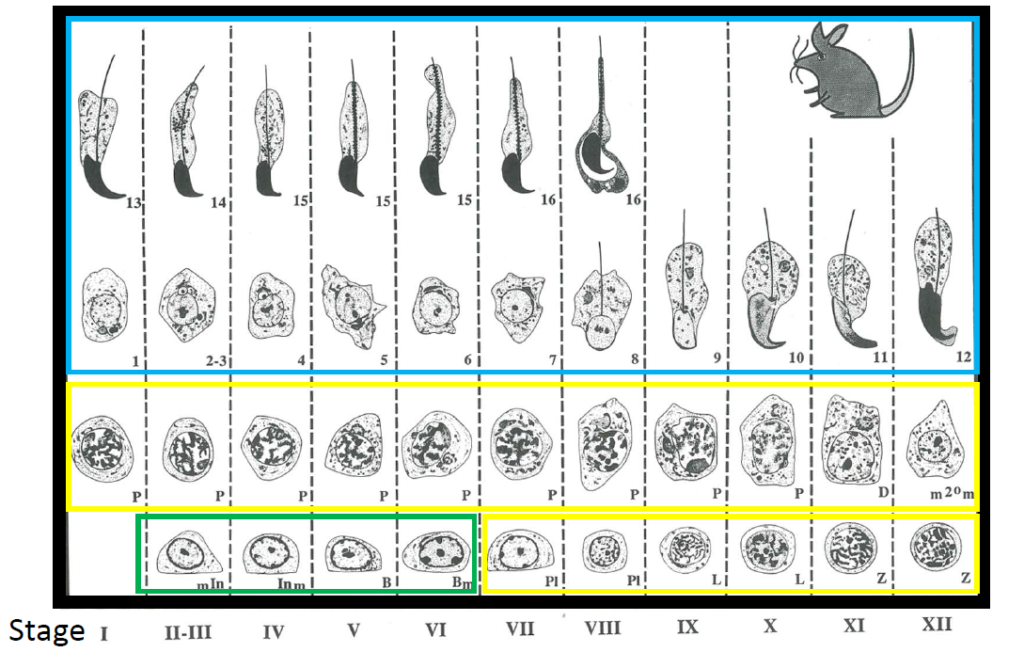
- 【コラム更新】精子がオタマジャクシ状になる理由を求めて 嶋田圭祐先生(酪農学園大学 獣医学群 獣医学類)
- 2026.05.22
- 【コラム更新】実験動物としての両生類を、いま改めて考える― イベリアトゲイモリとの出会いと、飼育・利用基盤の整備 ― 大林徹也先生(鳥取大学・研究推進機構)
- 2026.05.20
- 【コラム更新】完全長マウスゲノムの完成 廣瀬直毅先生(大阪大学 大学院医学系研究科)


米国獣医学会(AVMA)動物の安楽死指針2020年版出版記念 -紹介動画-
日本実験動物医学専門医協会は、AVMAと翻訳契約を取り交わし、「米国獣医学会 動物の安楽死指針(安楽死ガイドライン):2020年」版の翻訳本(翻訳者代表 黒澤努、鈴木真)を出版しました。本ガイドラインは、国際的に容認される具体的な安楽死法を示しており、主に獣医師を対象に記載されています。専門的ではありますが、最新の情報を網羅しており、獣医師以外の動物にかかわる方々の指針としても重要な文献です。(原文はこちら)
2013年度版から改訂された2020年版では、第3章にS1コンパニオンアニマル、S2実験動物、S3家畜、S4馬、S5鳥類、S6魚類と水生無脊椎動物、S7野生動物と7つの動物に区分されて記載されています。
日本実験動物医学会および日本実験動物医学専門医協会は、本指針が広く周知されることで、わが国の動物福祉がより向上することを期待します。また、実験動物ならびにその他の動物の人道的な取り扱いを広めるための啓蒙活動を継続していきます。
米国獣医学会(AVMA)動物の安楽死指針(安楽死ガイドライン)2020年版の紹介
炭酸ガスを用いた安楽死
Compassion Fatigue(共感疲労)
Compassion Fatigueについて、さらに知りたい方はこちらもご覧ください。

実験動物のリホーミング
実験動物の飼養及び保管並びに苦痛の軽減に関する基準では、第4章実験等の実施上の配慮の項において、「実験に供する期間をできるだけ短くする等実験終了の時期に配慮すること」と記されています。そして、実験動物の飼養及び保管並びに苦痛の軽減に関する基準の解説によると、実験計画の立案においては、「実験や術後観察の終了の時期(人道的エンドポイント)等について、具体的な計画を立案する必要がある。(p. 114)」と解説されています。また、人道的エンドポイントとは、「実験動物を激しい苦痛から解放するために実験を終了あるいは途中で中止する時期(すなわち安楽死処置を施す時期)を意味する。(p. 142)」と解説されています。こうしたことから、動物実験の終了とは、主として安楽死処置を施すこととも捉えられます。
一方で、安楽死処置については、上述の通り実験動物を激しい苦痛から解放するための措置である反面、「安全性に加え、安楽死処置実施者が感じる精神的不安、不快感、あるいは苦痛に配慮し、科学的研究の目的を損なわない限り、心理的負担の少ない安全な方法を選択すべきである。(p. 159)」とも解説されており、実施者にとっては精神的不安、不快感、あるいは苦痛といった心理的負担を伴う措置であるということも理解されています。
このような安楽死における実施者の心理的負担に関しては、「安楽死にまつわる諸問題」についてのコラムですでに紹介されていますが、動物実験が遂行される中で、必ずしも動物は苦痛を被って実験を終えるものでもありません。こうした動物に対してはどのようにエンドポイントを考えたらよいでしょうか。これらの動物にも安楽死処置を施すのでしょうか。その心理的負担は苦痛から解放するための安楽死処置の場合よりも大きいものになるかもしれません。他に選択肢はないのでしょうか。
- AVMA2020
- C57BL/6
- Compassion Fatigue
- ENU
- JALAM
- JALAM会員の寄稿
- JCLAM
- SPF
- イベント
- ウェビナー
- エンドポイント
- エンリッチメント
- くすり
- コットンラット
- コラーゲン
- サイエンスコミュニケーション
- シミュレーター
- スタニオカルシン-1
- ドライシッパー
- フォワードジェネティクス研究
- ブタ
- マウス
- ミネラル調節
- モデル動物
- リホーミング
- 代替法
- 前島賞
- 創薬研究
- 動物実験
- 動物福祉
- 動画
- 周術期管理
- 大学
- 学会
- 学術集会優秀発表賞
- 安全性
- 安楽死
- 実験動物
- 審査
- 情報発信
- 教育
- 教育委員会
- 文献紹介
- 海外
- 環境
- 筋ジストロフィー
- 系統
- 老化
- 蚊の退治法
- 食品検査
- 飼育室温度
- 飼育環境
- 麻酔

実験動物としての両生類を、いま改めて考える― イベリアトゲイモリとの出会いと、飼育・利用基盤の整備 ―
鳥取大学・研究推進機構
大林徹也
両生類というと、現在の生命科学では、再生や発生を専門とする一部の研究者が用いる特殊な動物、という印象をもたれやすいかもしれない。しかし歴史をたどると、両生類はむしろ、脊椎動物を用いた実験生物学のかなり早い時期から研究に使われてきた動物群である。イモリは古くから再生研究の対象となり、カエル類は発生学の材料として重要な役割を果たしてきた。イモリの再生現象は17世紀にはすでに実験生物学の対象となっており、イモリは最も長い歴史をもつ実験動物の一つと位置づけられている[1,2]。
両生類が基礎生物学だけでなく医学にも関わってきたことを示す分かりやすい例が、アフリカツメガエルである。1930年代には、妊婦の尿を用いた妊娠診断にツメガエルが使われていたことが知られている。妊婦尿中のホルモンに反応して排卵する性質を利用したもので、当時としては実用的な医学的検査法であった[3]。いまから見れば少し意外に感じられるかもしれないが、両生類は「基礎研究だけの動物」ではなく、かなり早い段階から医学とも接点をもっていたのである。この点を最初に確認しておくと、両生類がその後なぜ主流から外れていったのか、そして今なぜ見直す価値があるのかが見えやすくなる。
それにもかかわらず、20世紀後半以降、生命科学研究の中心は急速にマウスへ移っていった。もちろん、マウスが研究対象として優れていたことは間違いないが、より大きかったのは、近交系、安定供給、標準化された飼育条件、遺伝学的基盤、さらに分子生物学やゲノム情報の整備が、マウスでは非常に速く進んだことであろう[4,5]。一方、両生類では、面白い生命現象があることと、実験動物として使いやすいことが必ずしも一致していなかった。カエルと同様にイモリ研究には長い歴史がある一方で、飼育下繁殖、系統維持、標準化の面で難しさが残り、野外採集個体への依存も長く続いた[1,2]。その結果、両生類の利用が減ったというよりも、近代的な実験動物学が求めた条件への対応が、マウスのほうで先に進んだと考えるほうが実態に近いように思う。
私がイベリアトゲイモリと出会ったのは、2012年頃だったと思う。現在、広島大学両生類研究センター教授である林利憲先生から、「動物実験施設でイベリアトゲイモリを飼育したい」と相談されたことがきっかけであった。当時の私にとって、両生類は再生や発生の基礎研究で用いられる生き物という印象が強く、いわゆる実験動物という言葉とは少し距離があった。しかし林先生は、かなり早い時期から、イベリアトゲイモリが単に再生能力の高い有尾両生類というだけでなく、飼育下で安定して繁殖でき、将来的には遺伝学的解析にも使えるモデルになり得ることを見通しておられたのだと思う[1,2]。
イベリアトゲイモリは「大量繁殖できるイモリ」として、再生研究を分子・遺伝学レベルへと刷新するための必然的なモデル生物である[2]。イモリ研究は長い歴史をもち、レンズ再生に代表される古典的な再生現象で広く知られてきたが、繁殖が難しく、研究を大きく発展させにくいという弱点があった。ならば、大量繁殖できるイモリを導入すればよい、という発想である。この考え方はきわめて分かりやすい。しかもイベリアトゲイモリでは、四肢だけでなく、より複雑な器官の再生も研究対象となっている。たとえば心臓では、凍結損傷後の心筋組織が回復し、瘢痕組織の再構築や心筋細胞の増殖を伴う再生過程[6]が解析されている。また、生殖器官の面でも機能を失ったり除去されたりした精巣に代わって、新しい精巣が形成されるという特異な現象[7]がみられる。再生能力という古典的な魅力を保ちながら、実験動物として重要な繁殖性も備えているところに、本種の大きな価値がある。さらに近年は、非哺乳類による発生毒性試験にイベリアトゲイモリを用いる取り組み[8]も進んでおり、再生研究にとどまらない広がりを見せている。 実際に飼育に関わるようになると、イベリアトゲイモリは想像以上に「施設になじむ」動物であることが分かった。一般に両生類というと、特殊な環境を整えなければ維持できないような印象をもたれがちであるが、イベリアトゲイモリは、少なくとも実験室内での維持という点ではそうではない。林らは、プラスチック容器、簡易ろ過装置、水道水を基本とした低コストで簡便な飼育法を整え、雄は約6か月、雌は約9か月で性成熟し、1回150〜600個、年間では約5,000個にも及ぶ産卵が可能で、しかも通年産卵という大きな利点を示している[2]。私自身の感覚でも、実験用マウスを適切に管理できる施設環境があれば、イベリアトゲイモリの導入はそれほど難しくない。水棲動物としての配慮は必要であるものの、「マウス施設とはまったく別世界の生き物」ではないのである。

【図1:マウスの飼育ラックを改造したイベリアトゲイモリ飼育室システム】
この点は、イベリアトゲイモリ標準飼育プロトコルでもかなり具体的に示している[9]。室温は22〜26℃程度を基本とし、28℃以上の高温が数日続くことは避ける。照明は14時間明期・10時間暗期を基本とする。飼育水については、市販の魚類用フィルターろ過器を用いて緩やかにエアレーションを行うことで、水中の汚れの蓄積を抑え、水質を安定させやすくなる。一方で、イベリアトゲイモリの飼育においてエアレーションは必須ではなく、週3回程度の換水を行い、水質悪化を防ぐことができれば、無エアレーションでも飼育は可能である。重要なのは、エアレーションの有無そのものではなく、給餌量、個体密度、換水頻度を管理し、水質を悪化させないことである。幼生から幼体期にかけては、共食い防止のため低密度飼育を行い、底面積20×10 cm程度の容器に4〜6匹を目安とする。水深よりも底面積を重視し、水質悪化を避けるため、給餌と換水のタイミングをそろえる。幼生はブラインシュリンプを用い、変態期以降は配合飼料へ切り替える。変態期はとくに体表が脆弱で、水質悪化や感染の影響を受けやすいため、給餌内容と水環境の維持が重要である[9]。こうした条件を一つずつ整理していくと、イベリアトゲイモリは、両生類の中では比較的標準化しやすい種ではないかと感じる。

【図2:イベリアトゲイモリの幼生・幼体・成体】
コラム実験動物

完全長マウスゲノムの完成
大阪大学 大学院医学系研究科
廣瀬 直毅
生物の体の設計図は、A・T・G・Cの4種類の塩基の配列としてゲノムに刻まれています。ヒトゲノムの塩基配列の大部分は、今から約20年前に初めて解読1–3されました。そして2022年、米国の国立ヒトゲノム研究所、カリフォルニア大学サンタクルーズ校、ワシントン大学を中心とするテロメアtoテロメア・コンソーシアムにより、ついに短腕テロメアから長腕テロメアまでの完全長(telomere-to-telomere、T2T)ゲノムが、すべての常染色体とX染色体について解読されました4。さらに翌年にはY染色体の完全長ゲノムも解読5され、ヒトの完全長ゲノムの参照配列が完成しました。従来のヒトゲノムの参照配列にあった341箇所の塩基配列の未決定領域(「ギャップ」)も解消されたため、疾患の発症に関わるゲノム異常を探索する研究が加速すると期待されています。
ヒトゲノムの解読に並行して、医学・生物学の研究に供されているさまざまな実験動物のゲノムの塩基配列も解読されてきました。代表的な実験動物であるマウス(ハツカネズミ;Mus musculus)については、C57BL/6J系統のゲノムがマウスゲノム・シークエンシング・コンソーシアムによって2002年に初めて解読6されました。それ以降も追加解読が重ねられてきたものの、依然として塩基配列の未決定領域が約7,350万塩基対ほど残っていました。しかし2024年12月に中国のYu博士が率いる研究チームによってC57BL/6系統7、そして2025年11月に英国のKeane博士が率いる研究チームによってC57BL/6J系統およびCAST/EiJ系統の完全長ゲノムの解読結果が、それぞれ報告されました8。本稿では、まずこれらのマウスの完全長ゲノムについて概説したあと、克服された問題と背景にある技術革新にふれ、最後に将来の展望に言及します。
マウスの完全長ゲノムの解読
C57BL/6系統の完全長ゲノムは、半数体(haploid)の雄核発生性の胚性幹細胞(mouse haploid androgenetic embryonic stem cell、mhaESC)を使って解読されました7。この研究では19個の常染色体とX染色体の完全長ゲノムについて報告されましたが、Y染色体の完全長ゲノムの解析結果も2025年6月に公開されました。中国の研究チームによって解析されたこともあり、完全長ゲノムのデータは独自のウェブサイトにて維持・管理されています(データ保存場所)。
C57BL/6J系統とCAST/EiJ系統の完全長ゲノムは、オスのC57BL/6J系統とメスのCAST/EiJ系統を交配させたF1オス個体の胚性幹細胞(ES細胞)を用いて解読されました8。解読されたC57BL/6Jの完全長ゲノム(2.638 Gbp、約26億塩基対)は常染色体のみであり、X・Y染色体は未解読です。一方、解読されたCAST/EiJの完全長ゲノム(2.665 Gbp)は常染色体およびX染色体から構成され、Y染色体が未解読です。ミトコンドリアのゲノムはすでに解読されているため、今回新しく公開されたデータには含まれていません。完全長ゲノムの情報はC57BL/6J系統(GCA_964188535)とCAST/EiJ系統(GCA_964188545)それぞれについて公開されており、一般的なゲノムブラウザUCSC Genome Browserにて閲覧できます(C57BL/6J、CAST/EiJ)。さらにFASTAファイル(.fa)を扱えば、ゲノムの塩基配列をより詳細に検索できます(C57BL/6JとCAST/EiJそれぞれについてダウンロード可能)。また、The Allied Genetics Conferenceでの完全長ゲノムに関する講演の資料も、公開されています(英語資料)。ヒトの完全長ゲノムや他のさまざまな生物のゲノムのデータと同様に、C57BL/6J系統とCAST/EiJ系統の完全長ゲノムのデータは米国の国立生物工学情報センター(NCBI)のデータベースに集積されています。
(※リンク先は入稿した2026年4月末日現在にて有効)

ちゃんと向き合いたい、
実験動物のこと。
実験動物というとどんなイメージがあるでしょうか。
動物を実験に活用することへの抵抗感をお持ちの方もいらっしゃるかもしれません。
しかし、実験動物に携わる関係者の間では実験動物を科学的合理性だけでなく、
動物福祉の観点からも向き合い、飼育環境の改善、実験方法や規制の見直しといった工夫を
日々行っております。
当団体では、そういった日々進化する実験動物に関する情報を
様々なコンテンツを通じて発信しております。
当サイトが、実験動物に関心のある方々の理解を促進し、
よりよい動物と人間の共存関係を実現する一助となれば幸いに存じます。




米国獣医学会(AVMA)動物の安楽死指針2020年版出版記念 -紹介動画-
日本実験動物医学専門医協会は、AVMAと翻訳契約を取り交わし、「米国獣医学会 動物の安楽死指針(安楽死ガイドライン):2020年」版の翻訳本(翻訳者代表 黒澤努、鈴木真)を出版しました。本ガイドラインは、国際的に容認される具体的な安楽死法を示しており、主に獣医師を対象に記載されています。専門的ではありますが、最新の情報を網羅しており、獣医師以外の動物にかかわる方々の指針としても重要な文献です。(原文はこちら)
2013年度版から改訂された2020年版では、第3章にS1コンパニオンアニマル、S2実験動物、S3家畜、S4馬、S5鳥類、S6魚類と水生無脊椎動物、S7野生動物と7つの動物に区分されて記載されています。
日本実験動物医学会および日本実験動物医学専門医協会は、本指針が広く周知されることで、わが国の動物福祉がより向上することを期待します。また、実験動物ならびにその他の動物の人道的な取り扱いを広めるための啓蒙活動を継続していきます。
米国獣医学会(AVMA)動物の安楽死指針(安楽死ガイドライン)2020年版の紹介
炭酸ガスを用いた安楽死
Compassion Fatigue(共感疲労)
Compassion Fatigueについて、さらに知りたい方はこちらもご覧ください。
実験動物のリホーミング
実験動物の飼養及び保管並びに苦痛の軽減に関する基準では、第4章実験等の実施上の配慮の項において、「実験に供する期間をできるだけ短くする等実験終了の時期に配慮すること」と記されています。そして、実験動物の飼養及び保管並びに苦痛の軽減に関する基準の解説によると、実験計画の立案においては、「実験や術後観察の終了の時期(人道的エンドポイント)等について、具体的な計画を立案する必要がある。(p. 114)」と解説されています。また、人道的エンドポイントとは、「実験動物を激しい苦痛から解放するために実験を終了あるいは途中で中止する時期(すなわち安楽死処置を施す時期)を意味する。(p. 142)」と解説されています。こうしたことから、動物実験の終了とは、主として安楽死処置を施すこととも捉えられます。
一方で、安楽死処置については、上述の通り実験動物を激しい苦痛から解放するための措置である反面、「安全性に加え、安楽死処置実施者が感じる精神的不安、不快感、あるいは苦痛に配慮し、科学的研究の目的を損なわない限り、心理的負担の少ない安全な方法を選択すべきである。(p. 159)」とも解説されており、実施者にとっては精神的不安、不快感、あるいは苦痛といった心理的負担を伴う措置であるということも理解されています。
このような安楽死における実施者の心理的負担に関しては、「安楽死にまつわる諸問題」についてのコラムですでに紹介されていますが、動物実験が遂行される中で、必ずしも動物は苦痛を被って実験を終えるものでもありません。こうした動物に対してはどのようにエンドポイントを考えたらよいでしょうか。これらの動物にも安楽死処置を施すのでしょうか。その心理的負担は苦痛から解放するための安楽死処置の場合よりも大きいものになるかもしれません。他に選択肢はないのでしょうか。

JALAM会員提供動画
こちらでは当学会会員が提供する、実験動物・動物実験に関する動画を掲載していきます。
三種混合麻酔薬を用いた麻酔(マウス)
岡村匡史先生(国立国際医療研究センター研究所)
特集動画
実験動物としての両生類を、いま改めて考える― イベリアトゲイモリとの出会いと、飼育・利用基盤の整備 ―
鳥取大学・研究推進機構
大林徹也
両生類というと、現在の生命科学では、再生や発生を専門とする一部の研究者が用いる特殊な動物、という印象をもたれやすいかもしれない。しかし歴史をたどると、両生類はむしろ、脊椎動物を用いた実験生物学のかなり早い時期から研究に使われてきた動物群である。イモリは古くから再生研究の対象となり、カエル類は発生学の材料として重要な役割を果たしてきた。イモリの再生現象は17世紀にはすでに実験生物学の対象となっており、イモリは最も長い歴史をもつ実験動物の一つと位置づけられている[1,2]。
両生類が基礎生物学だけでなく医学にも関わってきたことを示す分かりやすい例が、アフリカツメガエルである。1930年代には、妊婦の尿を用いた妊娠診断にツメガエルが使われていたことが知られている。妊婦尿中のホルモンに反応して排卵する性質を利用したもので、当時としては実用的な医学的検査法であった[3]。いまから見れば少し意外に感じられるかもしれないが、両生類は「基礎研究だけの動物」ではなく、かなり早い段階から医学とも接点をもっていたのである。この点を最初に確認しておくと、両生類がその後なぜ主流から外れていったのか、そして今なぜ見直す価値があるのかが見えやすくなる。
それにもかかわらず、20世紀後半以降、生命科学研究の中心は急速にマウスへ移っていった。もちろん、マウスが研究対象として優れていたことは間違いないが、より大きかったのは、近交系、安定供給、標準化された飼育条件、遺伝学的基盤、さらに分子生物学やゲノム情報の整備が、マウスでは非常に速く進んだことであろう[4,5]。一方、両生類では、面白い生命現象があることと、実験動物として使いやすいことが必ずしも一致していなかった。カエルと同様にイモリ研究には長い歴史がある一方で、飼育下繁殖、系統維持、標準化の面で難しさが残り、野外採集個体への依存も長く続いた[1,2]。その結果、両生類の利用が減ったというよりも、近代的な実験動物学が求めた条件への対応が、マウスのほうで先に進んだと考えるほうが実態に近いように思う。
私がイベリアトゲイモリと出会ったのは、2012年頃だったと思う。現在、広島大学両生類研究センター教授である林利憲先生から、「動物実験施設でイベリアトゲイモリを飼育したい」と相談されたことがきっかけであった。当時の私にとって、両生類は再生や発生の基礎研究で用いられる生き物という印象が強く、いわゆる実験動物という言葉とは少し距離があった。しかし林先生は、かなり早い時期から、イベリアトゲイモリが単に再生能力の高い有尾両生類というだけでなく、飼育下で安定して繁殖でき、将来的には遺伝学的解析にも使えるモデルになり得ることを見通しておられたのだと思う[1,2]。
イベリアトゲイモリは「大量繁殖できるイモリ」として、再生研究を分子・遺伝学レベルへと刷新するための必然的なモデル生物である[2]。イモリ研究は長い歴史をもち、レンズ再生に代表される古典的な再生現象で広く知られてきたが、繁殖が難しく、研究を大きく発展させにくいという弱点があった。ならば、大量繁殖できるイモリを導入すればよい、という発想である。この考え方はきわめて分かりやすい。しかもイベリアトゲイモリでは、四肢だけでなく、より複雑な器官の再生も研究対象となっている。たとえば心臓では、凍結損傷後の心筋組織が回復し、瘢痕組織の再構築や心筋細胞の増殖を伴う再生過程[6]が解析されている。また、生殖器官の面でも機能を失ったり除去されたりした精巣に代わって、新しい精巣が形成されるという特異な現象[7]がみられる。再生能力という古典的な魅力を保ちながら、実験動物として重要な繁殖性も備えているところに、本種の大きな価値がある。さらに近年は、非哺乳類による発生毒性試験にイベリアトゲイモリを用いる取り組み[8]も進んでおり、再生研究にとどまらない広がりを見せている。 実際に飼育に関わるようになると、イベリアトゲイモリは想像以上に「施設になじむ」動物であることが分かった。一般に両生類というと、特殊な環境を整えなければ維持できないような印象をもたれがちであるが、イベリアトゲイモリは、少なくとも実験室内での維持という点ではそうではない。林らは、プラスチック容器、簡易ろ過装置、水道水を基本とした低コストで簡便な飼育法を整え、雄は約6か月、雌は約9か月で性成熟し、1回150〜600個、年間では約5,000個にも及ぶ産卵が可能で、しかも通年産卵という大きな利点を示している[2]。私自身の感覚でも、実験用マウスを適切に管理できる施設環境があれば、イベリアトゲイモリの導入はそれほど難しくない。水棲動物としての配慮は必要であるものの、「マウス施設とはまったく別世界の生き物」ではないのである。
【図1:マウスの飼育ラックを改造したイベリアトゲイモリ飼育室システム】
この点は、イベリアトゲイモリ標準飼育プロトコルでもかなり具体的に示している[9]。室温は22〜26℃程度を基本とし、28℃以上の高温が数日続くことは避ける。照明は14時間明期・10時間暗期を基本とする。飼育水については、市販の魚類用フィルターろ過器を用いて緩やかにエアレーションを行うことで、水中の汚れの蓄積を抑え、水質を安定させやすくなる。一方で、イベリアトゲイモリの飼育においてエアレーションは必須ではなく、週3回程度の換水を行い、水質悪化を防ぐことができれば、無エアレーションでも飼育は可能である。重要なのは、エアレーションの有無そのものではなく、給餌量、個体密度、換水頻度を管理し、水質を悪化させないことである。幼生から幼体期にかけては、共食い防止のため低密度飼育を行い、底面積20×10 cm程度の容器に4〜6匹を目安とする。水深よりも底面積を重視し、水質悪化を避けるため、給餌と換水のタイミングをそろえる。幼生はブラインシュリンプを用い、変態期以降は配合飼料へ切り替える。変態期はとくに体表が脆弱で、水質悪化や感染の影響を受けやすいため、給餌内容と水環境の維持が重要である[9]。こうした条件を一つずつ整理していくと、イベリアトゲイモリは、両生類の中では比較的標準化しやすい種ではないかと感じる。
【図2:イベリアトゲイモリの幼生・幼体・成体】
コラム実験動物
完全長マウスゲノムの完成
大阪大学 大学院医学系研究科
廣瀬 直毅
生物の体の設計図は、A・T・G・Cの4種類の塩基の配列としてゲノムに刻まれています。ヒトゲノムの塩基配列の大部分は、今から約20年前に初めて解読1–3されました。そして2022年、米国の国立ヒトゲノム研究所、カリフォルニア大学サンタクルーズ校、ワシントン大学を中心とするテロメアtoテロメア・コンソーシアムにより、ついに短腕テロメアから長腕テロメアまでの完全長(telomere-to-telomere、T2T)ゲノムが、すべての常染色体とX染色体について解読されました4。さらに翌年にはY染色体の完全長ゲノムも解読5され、ヒトの完全長ゲノムの参照配列が完成しました。従来のヒトゲノムの参照配列にあった341箇所の塩基配列の未決定領域(「ギャップ」)も解消されたため、疾患の発症に関わるゲノム異常を探索する研究が加速すると期待されています。
ヒトゲノムの解読に並行して、医学・生物学の研究に供されているさまざまな実験動物のゲノムの塩基配列も解読されてきました。代表的な実験動物であるマウス(ハツカネズミ;Mus musculus)については、C57BL/6J系統のゲノムがマウスゲノム・シークエンシング・コンソーシアムによって2002年に初めて解読6されました。それ以降も追加解読が重ねられてきたものの、依然として塩基配列の未決定領域が約7,350万塩基対ほど残っていました。しかし2024年12月に中国のYu博士が率いる研究チームによってC57BL/6系統7、そして2025年11月に英国のKeane博士が率いる研究チームによってC57BL/6J系統およびCAST/EiJ系統の完全長ゲノムの解読結果が、それぞれ報告されました8。本稿では、まずこれらのマウスの完全長ゲノムについて概説したあと、克服された問題と背景にある技術革新にふれ、最後に将来の展望に言及します。
マウスの完全長ゲノムの解読
C57BL/6系統の完全長ゲノムは、半数体(haploid)の雄核発生性の胚性幹細胞(mouse haploid androgenetic embryonic stem cell、mhaESC)を使って解読されました7。この研究では19個の常染色体とX染色体の完全長ゲノムについて報告されましたが、Y染色体の完全長ゲノムの解析結果も2025年6月に公開されました。中国の研究チームによって解析されたこともあり、完全長ゲノムのデータは独自のウェブサイトにて維持・管理されています(データ保存場所)。
C57BL/6J系統とCAST/EiJ系統の完全長ゲノムは、オスのC57BL/6J系統とメスのCAST/EiJ系統を交配させたF1オス個体の胚性幹細胞(ES細胞)を用いて解読されました8。解読されたC57BL/6Jの完全長ゲノム(2.638 Gbp、約26億塩基対)は常染色体のみであり、X・Y染色体は未解読です。一方、解読されたCAST/EiJの完全長ゲノム(2.665 Gbp)は常染色体およびX染色体から構成され、Y染色体が未解読です。ミトコンドリアのゲノムはすでに解読されているため、今回新しく公開されたデータには含まれていません。完全長ゲノムの情報はC57BL/6J系統(GCA_964188535)とCAST/EiJ系統(GCA_964188545)それぞれについて公開されており、一般的なゲノムブラウザUCSC Genome Browserにて閲覧できます(C57BL/6J、CAST/EiJ)。さらにFASTAファイル(.fa)を扱えば、ゲノムの塩基配列をより詳細に検索できます(C57BL/6JとCAST/EiJそれぞれについてダウンロード可能)。また、The Allied Genetics Conferenceでの完全長ゲノムに関する講演の資料も、公開されています(英語資料)。ヒトの完全長ゲノムや他のさまざまな生物のゲノムのデータと同様に、C57BL/6J系統とCAST/EiJ系統の完全長ゲノムのデータは米国の国立生物工学情報センター(NCBI)のデータベースに集積されています。
(※リンク先は入稿した2026年4月末日現在にて有効)